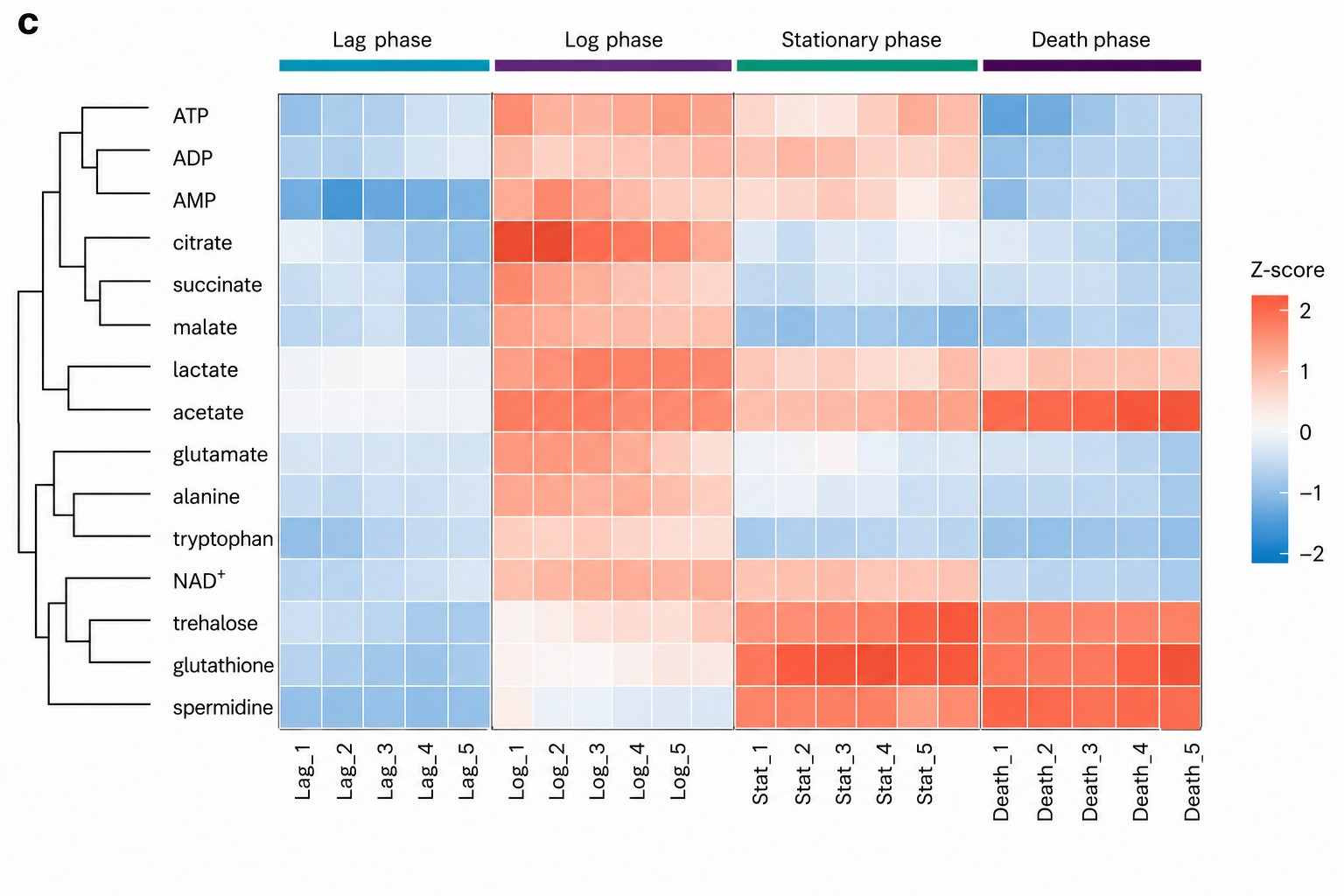
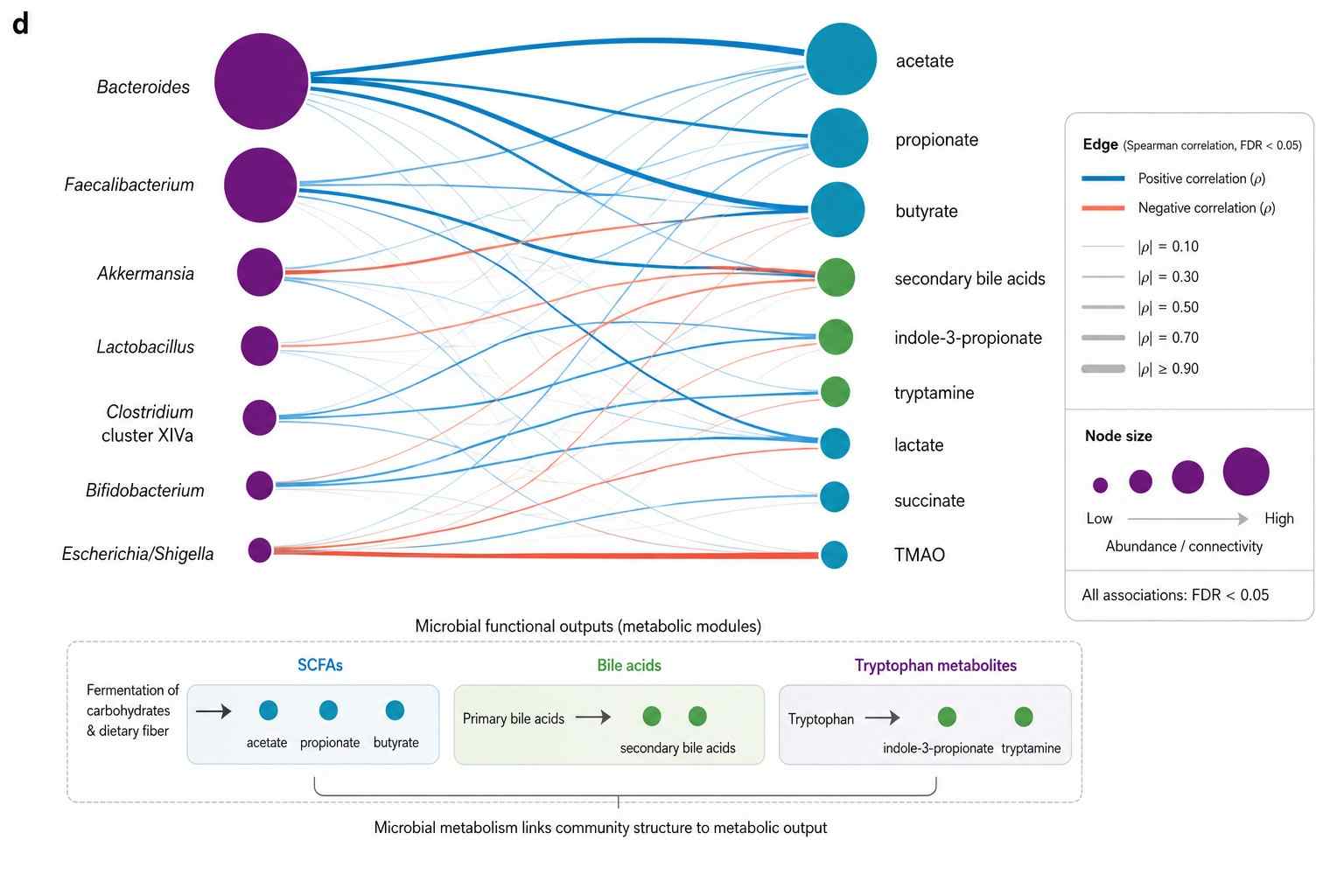
How We Analyze Microbial Metabolomes
A single bacterial culture contains volatile SCFAs at mM, polar central carbon intermediates at uM, non-polar lipids, and trace signaling molecules at nM — no single instrument covers this range. We layer four complementary approaches from the same sample, from broad discovery to targeted validation to multi-omics integration.
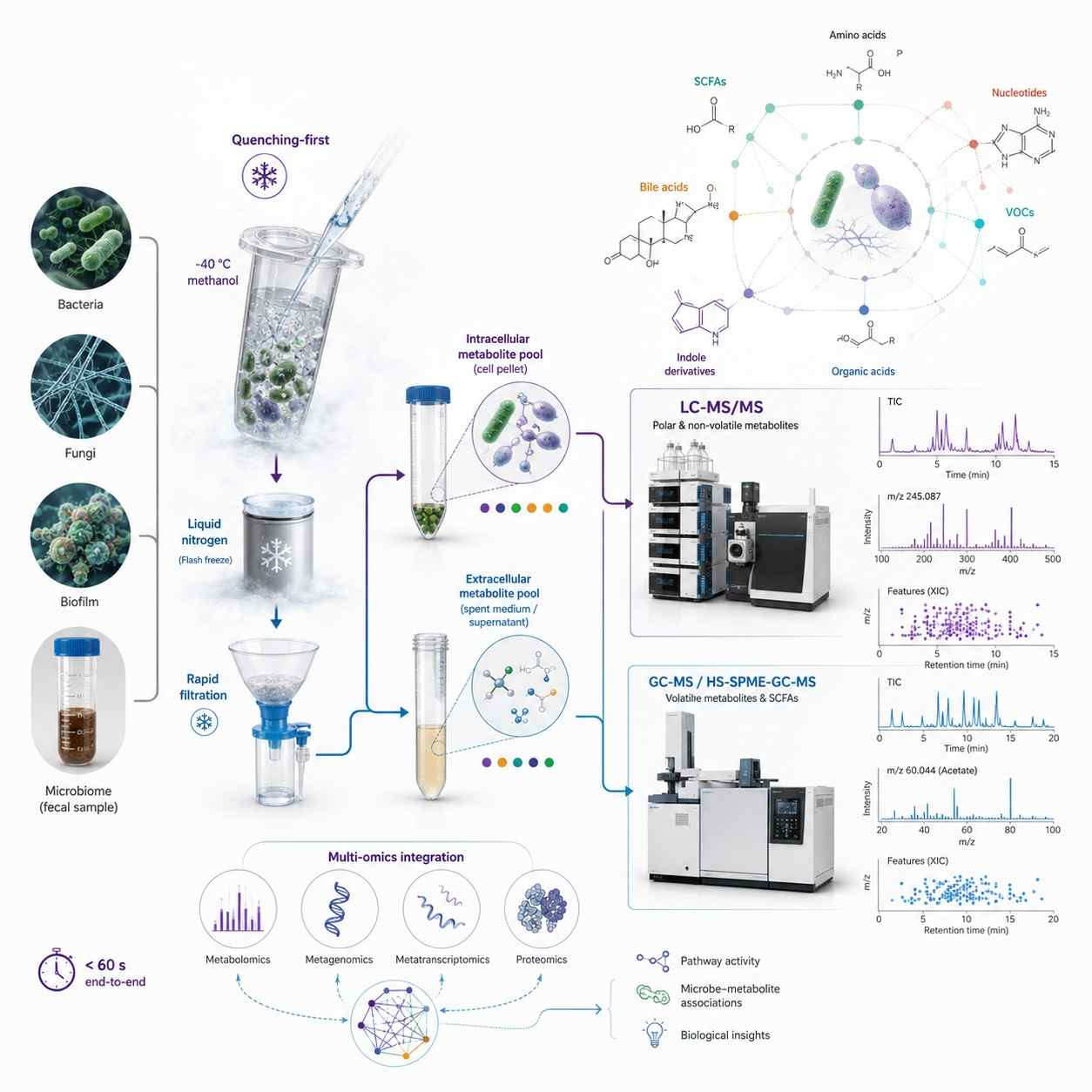
Metabolic Quenching & Sample Collection — Why It Matters for Microbial Metabolomics
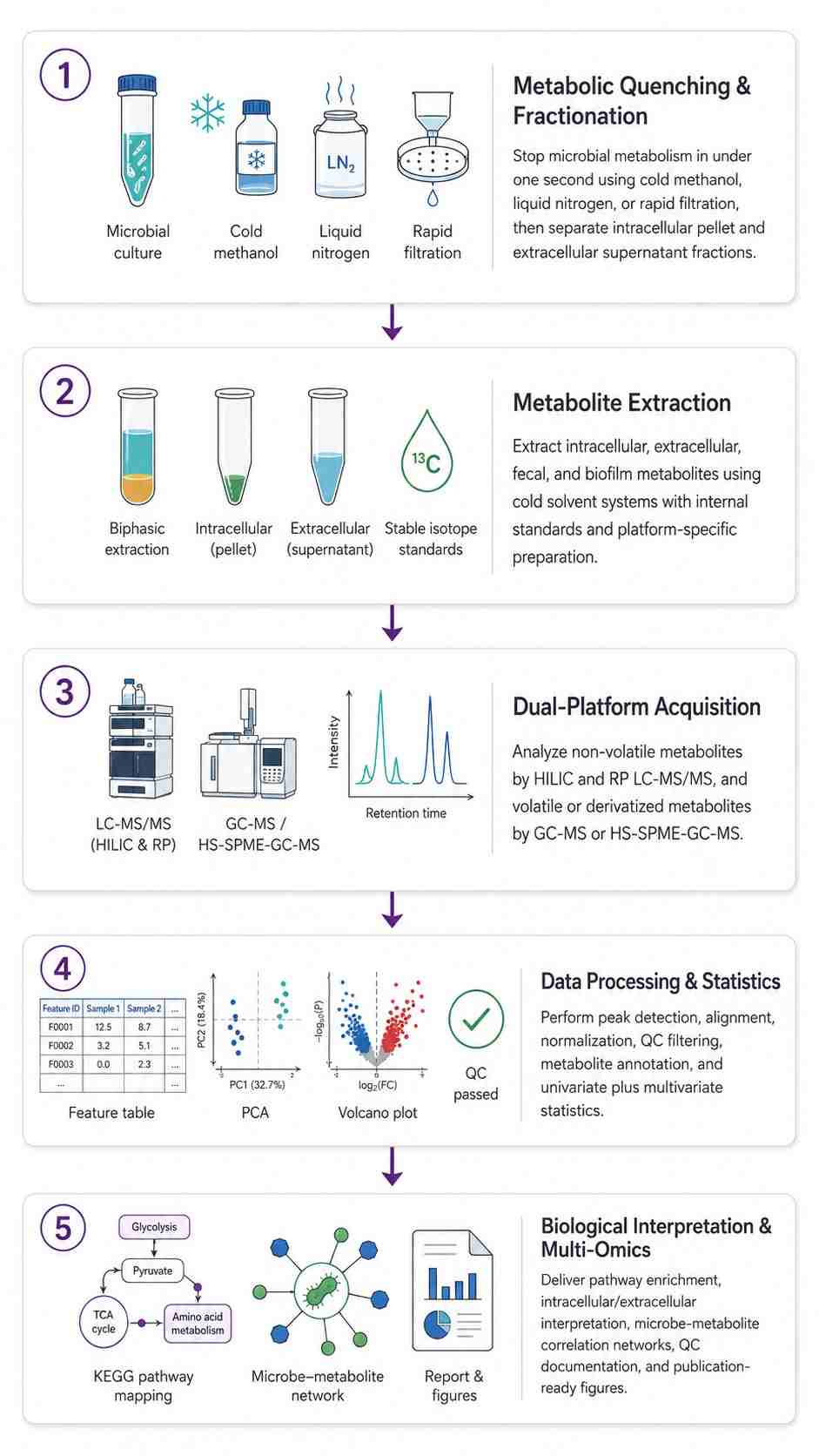
Unlike mammalian tissue where metabolism stops upon excision, a microbial sample remains metabolically active throughout collection and processing. A bacterial pellet centrifuged at room temperature for 10 minutes has a completely different metabolome than the same culture at the moment of harvest — ATP/ADP ratios collapse, central carbon intermediates shift, and volatile metabolites escape. These are the protocols we use to capture the true in vivo metabolome:
- Cold Methanol Quenching (-40 degree C) — Culture broth mixed with pre-chilled 60% methanol at 1:3-5 ratio, stopping all enzymatic activity in under 1 second. Cells pelleted at -20 degree C, washed, extracted. ATP and central carbon intermediates remain within 5% of in vivo values.
- Rapid Filtration + Liquid N2 — For filamentous fungi, biofilms, and environmental samples. Vacuum-filtered through 0.22 um membrane, washed with ice-cold PBS (under 5 sec), flash-frozen in liquid N2. Eliminates centrifugation — critical for organisms where metabolites leak during pelleting.
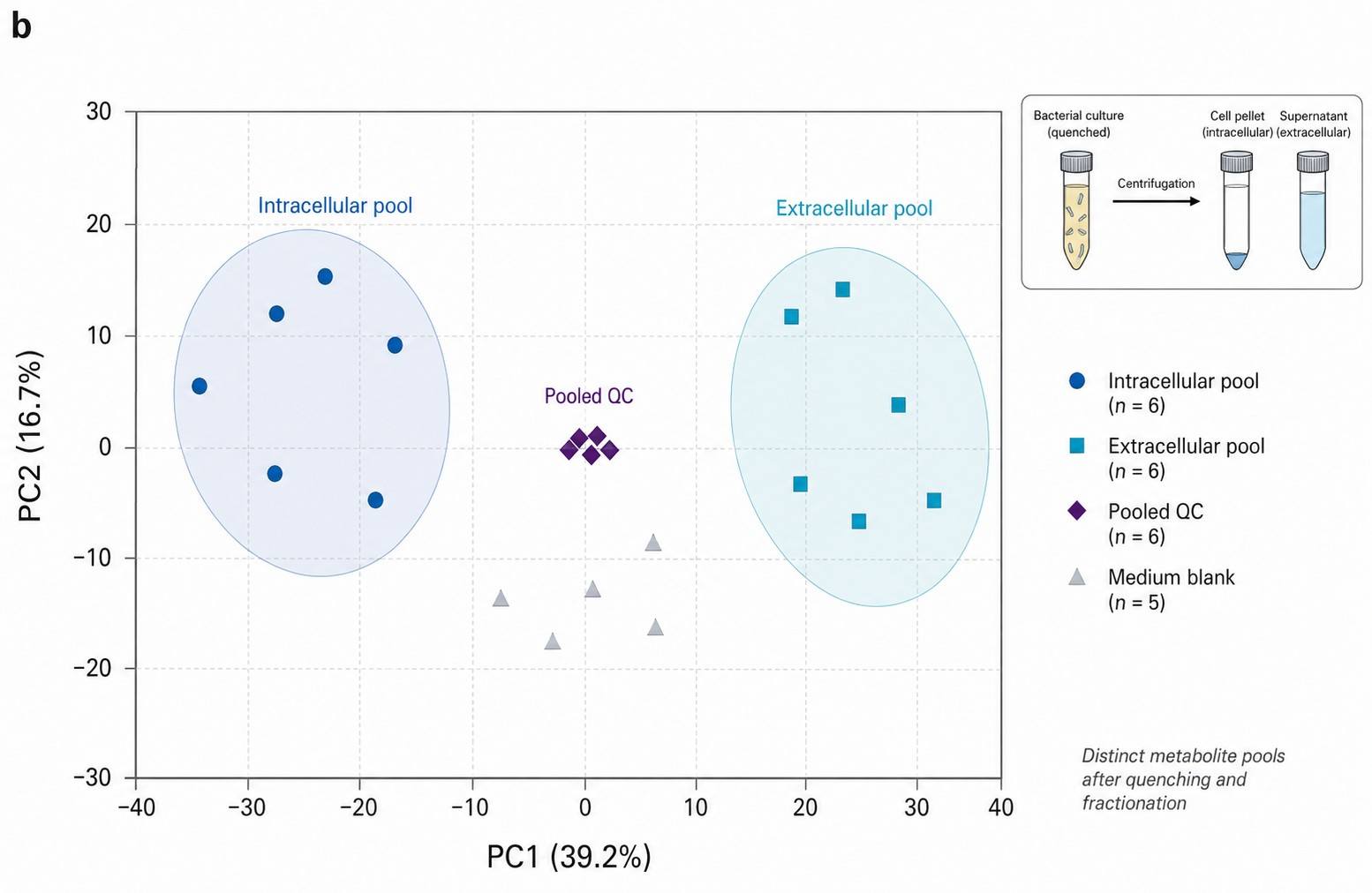
- Intracellular vs. Extracellular Separation — Quenched supernatant (exometabolome) and cell pellet (intracellular pool) extracted and analyzed independently. Distinguishes retained from secreted metabolites. Most CROs report a single value without this separation.
- Fecal & Complex Microbiome Samples — Flash-frozen in liquid N2 within 5 min of collection. Lyophilized for stable storage. Extracted with methanol:water (polar) and MTBE (lipids). Concentrations normalized to dry weight.
Analytical Platforms for Microbial Metabolomics
LC-MS/MS — Non-Volatile Metabolites
SCIEX QTRAP 6500+ with HILIC (polar metabolites: amino acids, nucleotides, central carbon intermediates, cofactors) and RP C18 (lipids, bile acids, non-polar metabolites) chromatography. Scheduled MRM for targeted quantification with stable isotope IS. Untargeted discovery on AB SCIEX TripleTOF 5600 (SWATH DIA, mass accuracy below 2 ppm) and Thermo Q Exactive Orbitrap (140,000 FWHM, HCD fragmentation).
GC-MS — Volatile & Derivatized Metabolites
Agilent 7890B-5977A GC-MS with EI source, DB-5MS/DB-FFAP columns. SIM for targeted quantification of SCFAs, organic acids, alcohols, and sugars (after derivatization). Thermo TSQ 9000 GC-MS/MS with SRM for low-abundance volatiles at sub-nM sensitivity. HS-SPME-GC-MS for microbial VOC profiling — headspace solid-phase microextraction captures volatile metabolites directly from culture headspace or fecal samples without solvent extraction.
Microbial Metabolomics Workflow — From Quenching to Biological Insight
Sample Types & Requirements for Microbial Metabolomics
| Sample Type |
Minimum Amount |
Quenching & Collection |
Storage & Shipping |
| Bacterial Culture (planktonic) |
1-5 x 10^7 cells per replicate (OD600 ~0.5-1.0, 10-50 mL culture) |
Cold methanol quenching (-40 degree C, 1:3-5 ratio). Pellet by centrifugation at -20 degree C, 5,000 x g, 5 min. Wash with cold methanol. Record growth phase, medium composition, temperature, and OD600. 5-6 biological replicates minimum. |
Pellet: -80 degree C, dry ice. Spent medium supernatant: collect separately, -80 degree C |
| Fungal Culture (mycelial) |
50-200 mg wet weight mycelium |
Rapid vacuum filtration through 0.22 um membrane, wash with ice-cold PBS (under 5 sec), flash-freeze filter + mycelium in liquid N2. Record growth phase and medium. |
-80 degree C, dry ice |
| Biofilm |
Scraped biomass from 1-3 wells (6-well plate) or equivalent surface area |
Scrape into ice-cold PBS, vacuum-filter, flash-freeze in liquid N2. Alternatively, cold methanol quenching directly on the growth surface. Record growth time and medium. |
-80 degree C, dry ice |
| Fecal / Stool (Human, Animal) |
100-200 mg fresh weight |
Collect into sterile cryovial, flash-freeze in liquid N2 within 5 min of defecation. Lyophilized aliquots recommended for long-term storage. Record diet, medication, and collection time. |
-80 degree C, dry ice |
| Fermentation Broth |
1-5 mL |
Centrifuge (5,000 x g, 5 min, 4 degree C). Collect supernatant (extracellular metabolites) and pellet (intracellular) separately. Flash-freeze both in liquid N2. Record fermentation time, substrate, pH, temperature. |
-80 degree C, dry ice |
| Soil / Sediment / Environmental |
5-10 g fresh weight |
Homogenize, remove debris. Flash-freeze in liquid N2 within 30 min of collection. Record soil type, moisture, pH, organic matter content, and collection depth. |
-80 degree C, dry ice |
Applications of Microbial Metabolomics
Specialized Microbial Research Services
The core workflow above — quenching, dual-platform LC-MS/MS + GC-MS, intra/extracellular separation — applies to any culturable microorganism: E. coli, Bacillus, Pseudomonas, Streptomyces, Saccharomyces, Aspergillus, and beyond. The services below add organism-specific quenching, targeted panels, and domain-specific data interpretation for projects requiring deeper expertise. Additional pathogen and species protocols available on request.
Microbial Metabolomics Deliverables
- Quantitative Data Tables — Absolute concentrations (uM, nmol/g dry weight, or nmol/10^6 cells) or normalized peak areas (untargeted) for all detected metabolites. Intra- and extracellular fractions reported separately. Excel and CSV.
- QC Report — Pooled QC RSD per metabolite class, IS recovery per sample, calibration curves (targeted), blank carryover, batch-effect documentation. Quenching efficiency indicators included.
- Metabolite Identification Table — MSI Level 1-4 confidence, m/z, RT, MS/MS matches, database IDs (HMDB, KEGG, METLIN, GNPS), and identification confidence per metabolite. Microbial-specific databases (MetaCyc, MiMeDB) searched in addition to standard libraries.
- Statistical Analysis & Figures — PCA/PLS-DA/OPLS-DA, volcano plots, hierarchical clustering heatmaps, KEGG/MetaCyc pathway enrichment, microbe-metabolite correlation networks (when paired with sequencing data). Publication-ready figures (300 DPI TIFF + vector PDF).
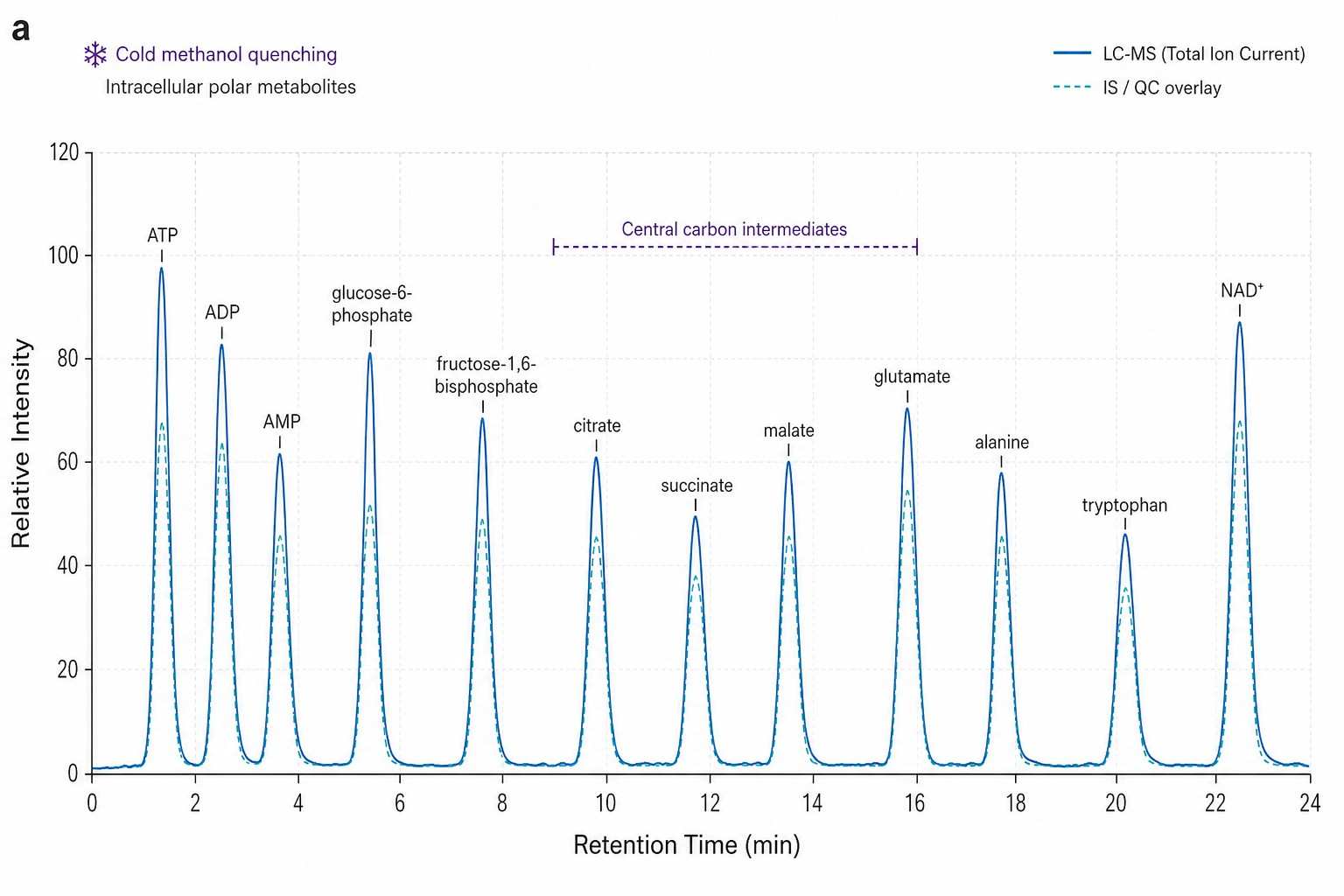
Microbial Metabolomics Data — Chromatograms, Multivariate Analysis & Pathway Maps
Case Study — How Metabolic Quenching Revealed the True Intracellular Metabolome of E. coli During Antibiotic Stress
Global metabolic profiling of Escherichia coli cultures: an evaluation of metabolite extraction methods for both extracellular and intracellular metabolites
Winder, C.L., Dunn, W.B., Schuler, S., Broadhurst, D., Jarvis, R., Stephens, G.M., & Goodacre, R. | Analytical Chemistry, 2008, 80, 2937-2945 | IF: 6.7
DOI: 10.1021/ac7023409
The Challenge
Bacterial metabolomics was plagued by a fundamental question no one wanted to address: how much of what you measure is actually inside the cell, and how much leaked out during sample processing? Cold methanol quenching — the standard method for stopping bacterial metabolism — causes membrane damage and metabolite leakage in many species. If 40% of your intracellular ATP leaks into the quenching solvent before you measure it, your "intracellular ATP concentration" is wrong. Winder et al. set out to quantify this leakage systematically — comparing six extraction methods across intracellular and extracellular fractions of E. coli with rigorous spike-recovery validation.
The Results
Using GC-MS untargeted metabolomics with cold methanol quenching followed by chloroform:methanol:water extraction (the same protocol we use for bacterial cultures), the study found:
- Cold methanol quenching at -48 degree C preserved the intracellular metabolome with minimal leakage (below 5% of total pool for most metabolites) — but only when quenching was completed within 1 second of harvest. Delayed quenching (even 30 seconds at room temperature) caused up to 40% loss of high-energy phosphates and a 3-fold increase in extracellular amino acids from cell lysis.
- Intracellular and extracellular fractions carried distinct metabolic signatures — 142 metabolites were detected in the intracellular fraction vs. 89 in the extracellular fraction, with only 38 shared. The two pools are metabolically distinct and must be analyzed separately.
- The method detected 200+ metabolite features per sample with intra-batch CV below 15% — demonstrating that rigorous quenching does not compromise data quality when executed correctly.
Why It Matters
This study confirmed what microbiologists suspected but couldn't quantify: metabolic quenching is not a technical detail — it is the difference between measuring the real metabolome and measuring an artifact. A bacterial culture processed without proper quenching reports "intracellular" metabolite concentrations that are contaminated with extracellular leakage, distorted by ongoing metabolism during centrifugation, and useless for modeling metabolic flux. Our quenching protocols are designed around this paper's findings: cold methanol at -40 degree C, 1:5 ratio, complete within 1 second, intra and extracellular fractions analyzed separately.
What This Means for You
If your experiment compares wild-type vs. mutant bacterial strains, antibiotic-treated vs. untreated cultures, or different growth phases — the quenching method determines whether you detect real metabolic differences or processing artifacts. Our protocol is validated to preserve the in vivo metabolome. Each sample's quenching parameters (temperature, ratio, time-to-quench) are documented in the QC report so you can confirm pre-analytical integrity.
How We Deliver the Same
- Cold methanol quenching at -40 degree C, 1:3-5 ratio, under 1 second — the validated protocol from Winder et al.
- Intracellular and extracellular fractions extracted and analyzed separately — not pooled into one ambiguous "cell pellet" value
- Stable isotope IS spiked at homogenization for absolute quantification with documented spike recovery
- Same GC-MS platform (Agilent 7890B-5977A) for metabolite detection across the same chemical diversity range
Reference
- Winder, C.L., Dunn, W.B., Schuler, S., Broadhurst, D., Jarvis, R., Stephens, G.M., & Goodacre, R. Global metabolic profiling of Escherichia coli cultures: an evaluation of metabolite extraction methods for both extracellular and intracellular metabolites. Analytical Chemistry 80, 2937-2945 (2008).
Selected Publications in Microbial Metabolomics
Global metabolic profiling of Escherichia coli cultures: an evaluation of metabolite extraction methods for both extracellular and intracellular metabolites
Winder, C.L., Dunn, W.B., Schuler, S., et al.
Journal: Analytical Chemistry
Year: 2008
DOI: https://doi.org/10.1021/ac7023409
microbeMASST: a taxonomically informed mass spectrometry search tool for microbial metabolomics data
Zuffa, S., Schmid, R., Bauermeister, A., et al.
Journal: Nature Microbiology
Year: 2024
DOI: https://doi.org/10.1038/s41564-023-01575-9
A phylogeny-guided metabolomics approach links specialized metabolism to bacterial ecology
Gavriilidou, A., Kautsar, S.A., Zaburannyi, N., et al.
Journal: Nature Microbiology
Year: 2024
DOI: https://doi.org/10.1038/s41564-024-01645-6
High-coverage metabolomics uncovers microbiota-driven biochemical landscape of interorgan transport and gut-brain communication in mice
Lai, Y.J., Liu, Y.L., Tsai, M.L., et al.
Journal: Nature Communications
Year: 2021
DOI: https://doi.org/10.1038/s41467-021-26508-2
B cell-intrinsic epigenetic modulation of antibody responses by dietary fiber-derived short-chain fatty acids
Sanchez, H.N., Moroney, J.B., Gan, H., et al.
Journal: Nature Communications
Year: 2020
DOI: https://doi.org/10.1038/s41467-019-13603-6
The olfactory receptor Olfr78 promotes differentiation of enterochromaffin cells in the mouse colon
Dinsart, G., Leprovots, M., Lefort, A., et al.
Journal: EMBO Reports
Year: 2024
DOI: https://doi.org/10.1038/s44319-023-00013-5
Microbial dysbiosis associated with impaired intestinal Na+/H+ exchange accelerates and exacerbates colitis in ex-germ free mice
Harrison, C.A., et al.
Journal: Mucosal Immunology
Year: 2018
DOI: https://doi.org/10.1038/s41385-018-0035-2
Pregnancy specific shifts in the maternal microbiome and metabolome in the BPH5 mouse model of superimposed preeclampsia
Beckers, K.F., et al.
Journal: PLOS ONE
Year: 2024
DOI: https://doi.org/10.1371/journal.pone.0287145