Plant Hormone Signaling & Targeted Quantification
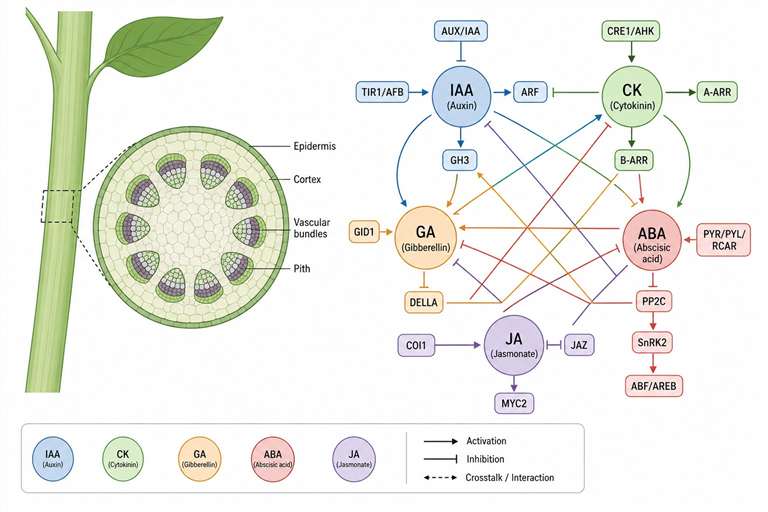
Plant hormones (phytohormones) are a structurally diverse set of signaling molecules that orchestrate virtually every aspect of plant life—from seed germination, root architecture, and shoot development to flowering, fruit ripening, and stress adaptation. These compounds operate at trace concentrations (pmol–fmol per gram fresh weight) through complex synergistic and antagonistic networks, making their accurate quantification both critically important and analytically challenging.
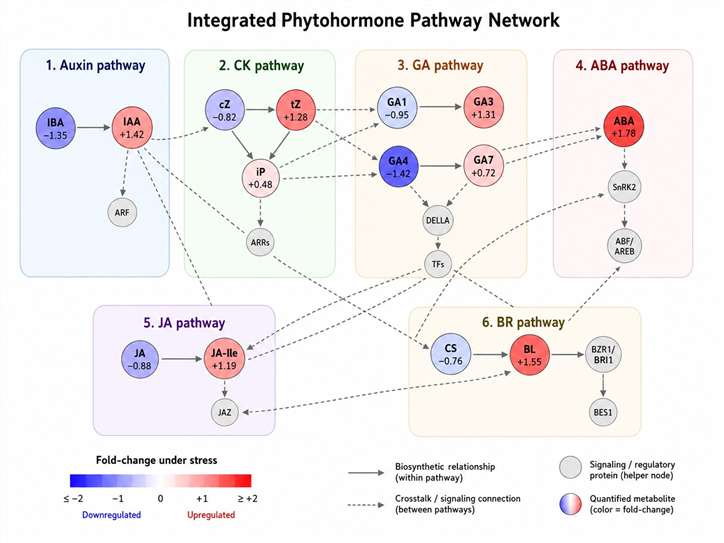
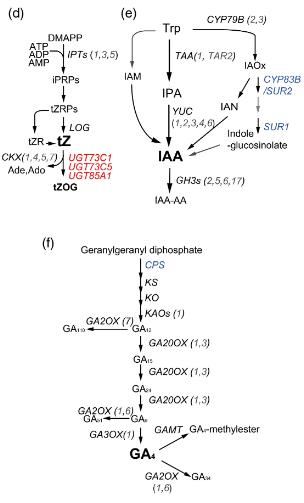
The major hormone families—auxins, cytokinins (CKs), gibberellins (GAs), abscisic acid (ABA), ethylene (ET), jasmonates (JAs), salicylic acid (SA), brassinosteroids (BRs), and strigolactones (SLs)—each regulate distinct but interwoven processes. For example, the auxin–cytokinin balance determines shoot vs. root architecture, ABA drives stomatal closure under drought, and the JA/ET module orchestrates defense against necrotrophic pathogens.
Targeted phytohormone panel metabolomics via LC–MS/MS and GC–MS/MS provides the most reliable approach for accurate hormone quantification at the pathway level. Unlike untargeted approaches, which survey all detectable signals and may miss low-abundance regulators, targeted MRM-based methods achieve the attomole-level sensitivity and structural specificity required for trace hormones. This enables you to:
(i) measure absolute hormone pool sizes and key metabolic ratios (e.g., IAA/CK, ABA/JA, GA/BR) that define developmental and stress states
(ii) monitor specific regulatory nodes such as auxin signaling, ABA/JA/ET stress cascades, GA/BR growth regulation, strigolactone/karrikin branching control, and peptide hormone signaling pathways
(iii) compare across plant tissues, developmental stages, stress treatments, and genetically modified lines with batch-to-batch data comparability.
Key Challenges in Targeted Phytohormone Analysis
Phytohormone analysis presents a unique set of analytical challenges not encountered in routine metabolomics. Low endogenous concentrations, severe matrix interference from pigments and polyphenols, and the need to cover multiple chemically distinct classes in a single run all contribute to the difficulty. Creative Proteomics addresses each barrier with targeted solutions:
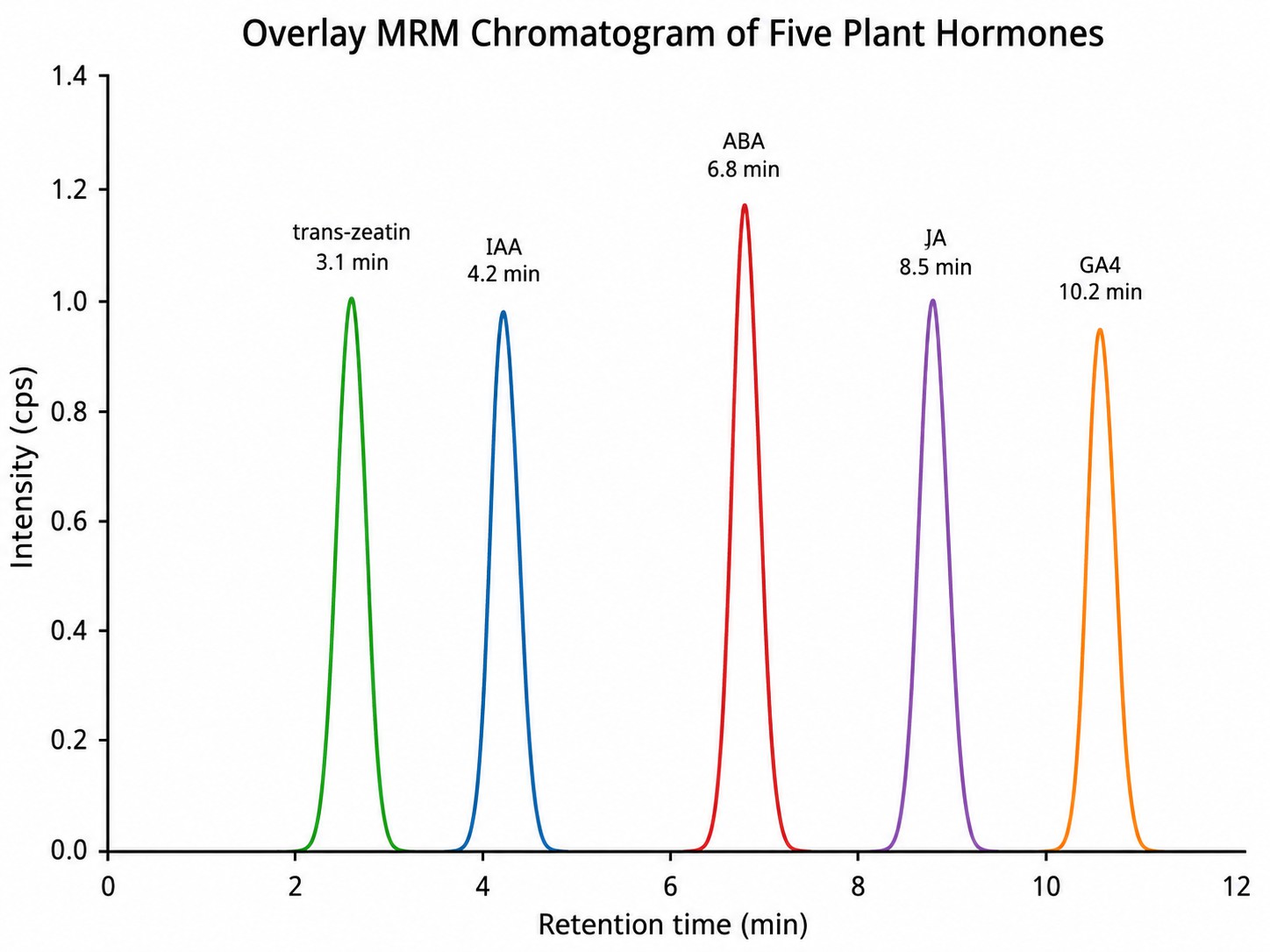
- Low abundance across structurally diverse classes: Phytohormones span a wide polarity range—from hydrophobic BRs to highly polar CK nucleotides—and exist at sub-ppm levels. Our workflow combines class-specific SPE enrichment and multi-mode LC (RP–C18 for most classes; HILIC for polar CKs and precursors) to maximize recovery across the full chemical spectrum. MRM acquisition on triple-quadrupole platforms (SCIEX QTRAP 6500+, Agilent 6495C) delivers attomole-level LODs.
- Matrix-driven ion suppression: Pigmented leaf tissue, resinous seed coats, and woody roots introduce co-eluting compounds that suppress ionization. We apply two-step SPE cleanup (mixed-mode anion exchange + RP) and matrix-matched calibration to correct for suppression, plus up to 30 isotope-labeled internal standards (D6-ABA, D5-tZ, D2-IAA, D6-SA, D2-JA, D2-GA4, D3-DHZ, and others) for per-analyte normalization.
- Isomer and epimer resolution: Many hormone classes contain structural and stereoisomers with distinct biological activities—cis- vs. trans-zeatin, GA1 vs. GA3, D/L-amino acid conjugates. We deploy chiral LC phases and optimized gradient programs to achieve baseline separation of critical isomers, supported by authenticated standard libraries for each target.
- Cross-class coverage in a single analytical run: Comprehensive hormonomics requires simultaneous quantification of compounds with vastly different physicochemical properties. Our integrated method uses time-segmented MRM scheduling, polarity switching, and derivatization strategies (dansyl chloride for BRs; MSTFA for GAs by GC–MS) to capture 100+ analytes from 10+ hormone classes in a single injection sequence.
- Labile compound preservation: GAs, BRs, and certain CK phosphates are prone to degradation during extraction and storage. We employ flash-freeze harvest protocols, cold methanolic extraction with antioxidant stabilization (BHT, ascorbic acid), and minimized sample handling to preserve labile targets.
Phytohormone Detection Panels & Pathway Coverage
Creative Proteomics offers a suite of LC–MS/MS and GC–MS/MS-based targeted panels covering major phytohormone classes, pathway-specific intermediates, and signaling metabolites. Panels are pre-configured for rapid deployment or fully customizable to match your species, tissue, and research question.
Standard Quantification Panels
| Panel |
Included Analytes |
| Core Phytohormone Panel (20+ targets) |
Indole-3-acetic acid (IAA), Indole-3-butyric acid (IBA), ABA, phaseic acid (PA), jasmonic acid (JA), jasmonoyl-isoleucine (JA-Ile), methyl jasmonate (MeJA), salicylic acid (SA), trans-zeatin (tZ), cis-zeatin (cZ), isopentenyladenine (iP), isopentenyladenosine (iPR), dihydrozeatin (DZ), GA1, GA3, GA4, GA7, brassinolide (BL), castasterone (CS), ethylene (via ACC) |
| Comprehensive Phytohormone Panel (100+ targets) |
All core phytohormones plus:
GA5, GA6, GA8, GA9, GA13, GA14, GA15, GA19, GA20, GA24, GA29, GA44, GA51, GA53, 6-deoxocastasterone (6-deoxoCS), typhasterol (TY), strigol, orobanchol, 5-deoxystrigol (5-DS), carlactonoic acid, 12-hydroxyjasmonic acid (12-OH-JA), salicylic acid 2-O-β-D-glucoside (SAG), zeatin riboside (ZR), 1-naphthaleneacetic acid (NAA), conjugated forms, and phase II metabolites. |
Pathway-Focused Panels
| Panel |
Key Targets / Pathways |
| Auxin Signaling & Metabolism |
IAA, IBA, NAA, IAA-Asp, IAA-Glu, IAA-Ala, tryptophan (precursor), indole-3-pyruvate (IPyA), indole-3-acetaldehyde (IAAld) |
| ABA/JA/ET Stress Response |
ABA, PA, DPA, JA, JA-Ile, MeJA, 12-OH-JA, SA, SAG, ACC (ethylene precursor), phaseic acid reductase products |
| CK & BR Growth Regulation |
tZ, cZ, iP, iPR, BAP, DZ, ZR, BL, CS, 6-deoxoCS, TY, 28-homobrassinolide |
| Gibberellin Profiling |
GA1–GA53 panel covering early C-13 hydroxylation and non-hydroxylation pathways, bioactive GAs (GA1, GA3, GA4, GA7), and catabolites; GC–MS/MS option for enhanced GA resolution |
| Strigolactone & Karrikin |
Strigol, orobanchol, 5-deoxystrigol (5-DS), carlactonoic acid, GR24 (internal standard), karrikin KAR1 |
Functional Biomarker Panels
| Panel |
Representative Classes |
| Peptide Hormones & Signaling Peptides |
Phytosulfokine (PSK), systemin, CLV3, RALF, CLE peptides, IDA, EPF (customized on request) |
| Polyamines & Nitric Oxide Metabolites |
Putrescine, spermidine, spermine, NO metabolites (nitrate, nitrite, S-nitrosothiols) |
| Melatonin & Related Indoles |
Melatonin, serotonin, tryptamine, 5-hydroxyindole-3-acetic acid (5-HIAA) |
| Oxylipins & Defense Signals |
12-OPDA, dinor-OPDA, 9-HOT, 13-HOT, 9-KOT, 13-KOT, and other oxylipin intermediates |
Customization & Add-ons
| Category |
Options |
| Target List |
Define your own list from phytohormones, conjugated forms, precursors, and custom signaling metabolites |
| Chiral / Isomer Resolution |
Support for cis/trans-zeatin differentiation, D/L isomer separation, and GA epimer resolution using chiral stationary phases or derivatization strategies |
| Internal Standards |
D-/13C-labeled internal standards for each hormone class (absolute quantification or semi-quantitative screening) |
| Labile Compound Stabilization |
Specialized workflows for GAs, BRs, and peptide hormones prone to degradation during extraction and storage |
| Multi-Omics Integration |
Parallel transcriptomics, proteomics, or untargeted metabolomics from the same biological sample for network-level analysis |
Why Choose Our Phytohormone Analysis Service?
- Curated Library of 150+ Phytohormones & Signaling Metabolites
Our in-house MS/MS library covers auxins, CKs, GAs, ABA, BRs, JAs, SA, SLs, peptide hormones, oxylipins, and related conjugates—enabling rapid panel configuration, confident annotation, and seamless expansion across interconnected pathways.
- Up to 30 Isotope-Labeled Internal Standards
We deploy hormone-specific stable isotope-labeled internal standards (D6-ABA, D5-tZ, D2-IAA, D6-SA, D2-JA, D2-GA4, D3-DHZ, 13C6-IAA, and others) to correct for matrix effects, extraction recovery, and ionization efficiency—ensuring absolute quantification accuracy across structurally diverse targets.
- Advanced Matrix Cleanup for Complex Tissues
Data undergo two-step SPE cleanup, isobaric interference correction, ion ratio QC, and manual peak review by experienced mass spectrometrists—ensuring data integrity even in challenging matrices such as pigmented leaf tissue, woody roots, resinous seed coats, and senescent material.
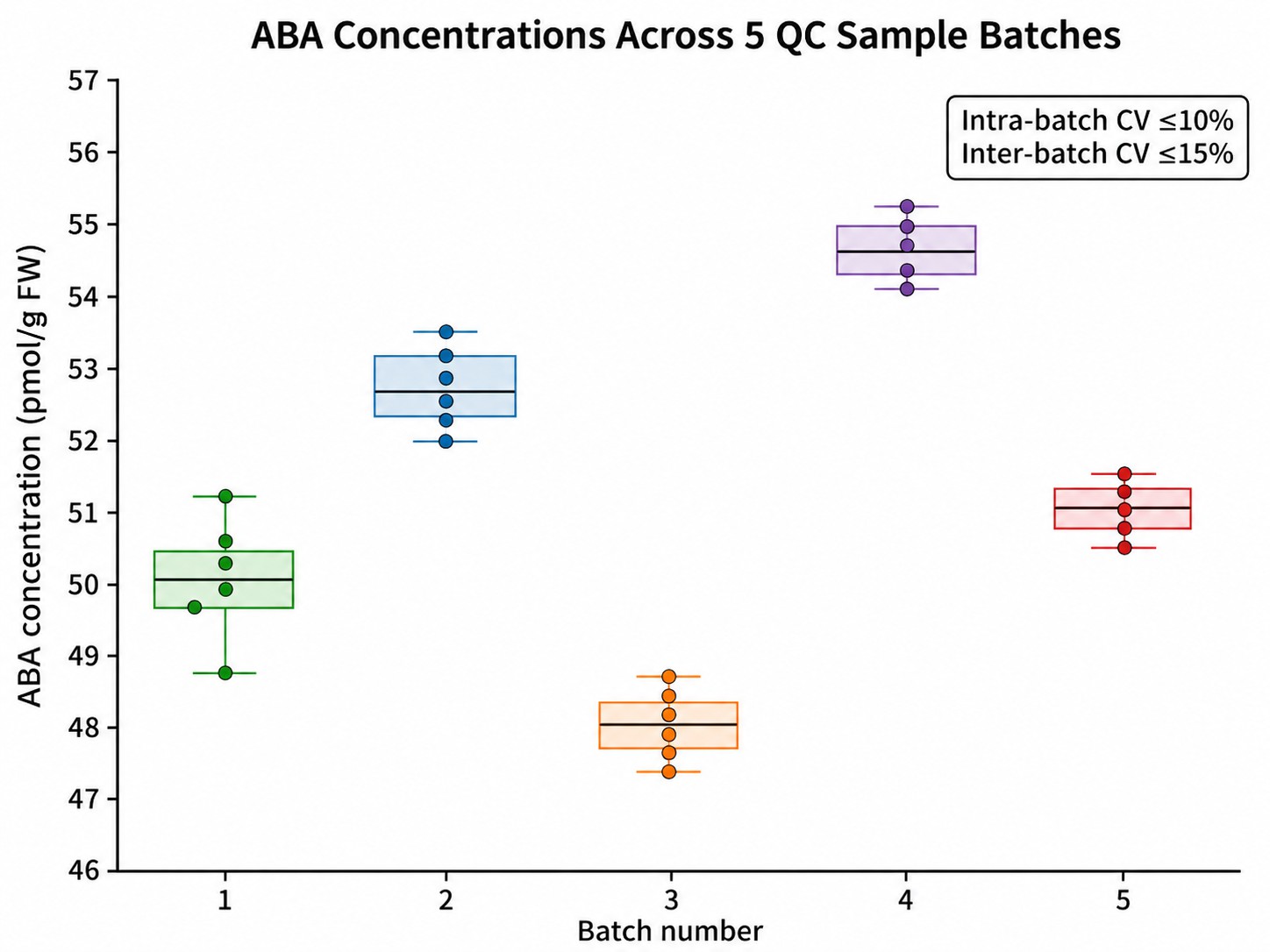
- Batch-Reproducible & Method-Locked Workflows
Ideal for multi-batch studies, long-term developmental time courses, and multi-site comparisons. Methods are locked per project; inter-batch bridging samples and system suitability benchmarks ensure data comparability across weeks or months of acquisition.
LC–MS/MS Platform & Method Performance
Analytical Platform
LC–MS/MS (Primary Platform)
Mass Spectrometer: Triple Quadrupole MS (SCIEX QTRAP series)
Ionization Mode: Electrospray Ionization (ESI), Positive/Negative polarity switching
LC System: UHPLC or HPLC (depending on resolution and throughput requirements)
Chromatography: Reversed Phase C18 (most classes); HILIC (polar CKs and precursors); Chiral phases (isomer separation)
Derivatization Options: Dansyl chloride (BRs), MSTFA (GAs by GC–MS), or derivatization-free (native) based on analyte class
GC–MS/MS (Optional / Confirmatory)
Available for orthogonal confirmation of volatile derivatives (GAs, ABA, ACC) with EI-MS library matching
Method Performance
| Parameter |
Typical Range |
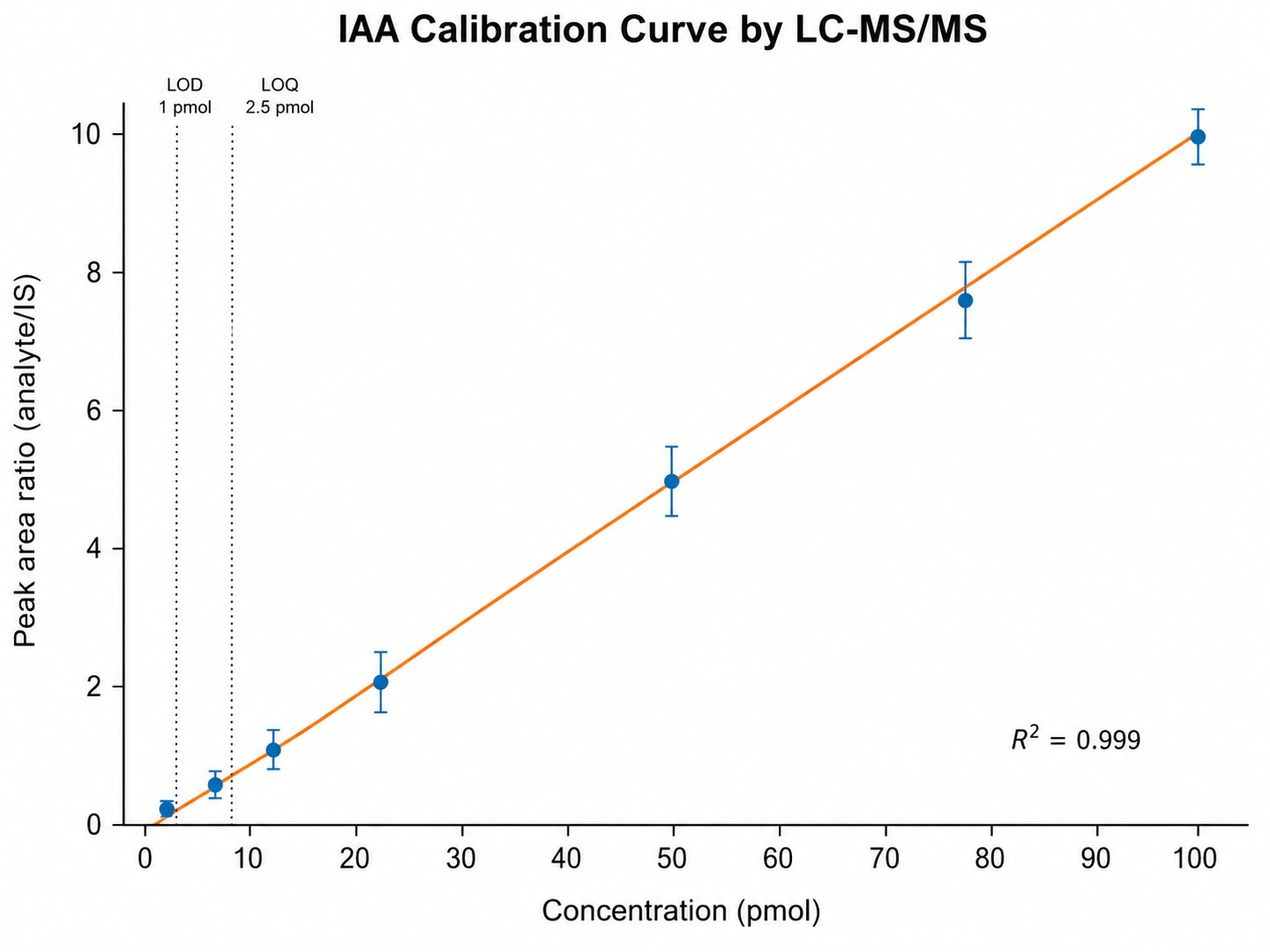
| LOD / LOQ |
0.01–1 pmol (on-column), analyte-dependent |
| Dynamic Range |
4–5 orders of magnitude |
| Quantification Type |
Absolute (isotope dilution) and/or relative (normalized to internal standards) |
| Precision (CV%) |
≤ 10% intra-batch; ≤ 15% inter-batch |
| Accuracy (Spike Recovery) |
85–115% |
| Injection Volume |
Typically 2–10 μL |
Internal Standards & Calibration
- Internal Standards: Up to 30 stable isotope-labeled phytohormones and pathway-specific analogs
- Calibration Strategy: 6–8 point standard curves, matrix-matched or surrogate matrix as appropriate
- System Suitability: Ion ratio QC, retention time window verification, peak shape assessment, carryover checks, and blank subtraction per batch
Phytohormone Analysis Workflow
Sample Submission Guide for Phytohormone Analysis
| Sample Type |
Amount Required |
Preparation Instructions |
Storage & Shipping |
| Leaf / Shoot |
10–50 mg FW |
Snap-freeze in liquid N₂ within 30 sec of excision; cryogenic grinding to fine powder; avoid pigment-rich veins if targeting low-abundance GAs/BRs |
−80°C; ship on dry ice |
| Root |
20–100 mg FW |
Wash gently in ice-cold water to remove soil, blot dry, snap-freeze immediately. Avoid prolonged washing (>2 min) to prevent hormone leakage |
−80°C; ship on dry ice |
| Seed / Grain |
50–200 mg FW |
Remove seed coat if needed; homogenize in liquid N₂ or cold 80% methanol. Stored seeds may have low ABA/IAA levels, requiring higher sample input |
−80°C; ship on dry ice |
| Fruit / Flower |
50–200 mg FW |
Remove non-target tissues (skin, pit, sepals); homogenize in liquid N₂; aliquot to avoid freeze–thaw. Ethylene-rich samples require ACC quantification route |
−80°C; ship on dry ice |
| Cell Culture / Callus |
1 × 10⁶ cells or 100 mg |
Vacuum-filter or centrifuge to remove medium; wash with cold PBS; snap-freeze pellet. Include medium blank to correct for secreted hormones |
−80°C; ship on dry ice |
| LCM / Microdissected Cells |
≥ 1,000 cells |
Laser-capture microdissection directly into ice-cold lysis buffer; minimize handling time to preserve labile SLs and BRs |
−80°C; ship on dry ice |
Notes
- Avoid repeated freeze-thaw cycles—plant hormones are highly labile
- Collect all samples at the same time of day (circadian hormone variation can exceed 2-fold for IAA and ABA)
- Clearly label each tube with sample ID, tissue type, developmental stage, date and time of collection
- Submit sample list (Excel preferred) with matching metadata including treatment conditions
- Contact us before sending rare, low-volume, or highly pigmented matrices for protocol optimization
Deliverables: What You Receive from Phytohormone Analysis
- Quantification tables: absolute (pmol/g FW) and/or relative hormone concentrations (.csv / Excel)
- Calibration & QC report: calibration curves, accuracy, precision, recovery, carryover checks per hormone class
- Raw & processed data: instrument raw files, processed peak lists, MRM chromatograms
- Method documentation: LC–MS/MS or GC–MS/MS parameters, derivatization protocol, internal standard setup
- Pathway annotations: KEGG / HMDB / CAS IDs linked to quantified phytohormones
- Final summary report: sample notes, QC results, interpretation highlights
Applications of Phytohormone Profiling
Our phytohormone profiling service supports researchers and breeders across plant science disciplines—from model species to crop plants and medicinal herbs. Applications span fundamental developmental biology, stress physiology, agricultural biotechnology, and multi-omics integration.
Integrated Transcriptome and Hormone Profiling Reveals Dynamic Responses to Moderate Dehydration Stress in Arabidopsis
Background
Plant responses to dehydration stress involve complex molecular and hormonal changes coordinated across multiple timescales. While the role of ABA in drought response is well established, the temporal dynamics of the full hormone network—including JA, SA, IAA, CKs, and GAs—during moderate dehydration remained poorly characterized. Single-hormone measurements or single time-point analyses fail to capture the regulatory crosstalk that drives adaptive responses.
Challenge: Characterize the temporal dynamics of genome-wide gene expression and comprehensive hormone profiles during moderate dehydration stress in Arabidopsis, with a focus on hormone–transcriptome integration to identify regulatory hubs and crosstalk mechanisms.
Key Findings (from the published study)
- Bi-phasic ABA accumulation observed during moderate dehydration: an early rapid increase (stomatal closure phase, 1–6 h) followed by sustained accumulation (protective gene induction phase, 12–72 h)
- Dynamic transcriptome reprogramming with distinct early (0–6 h) and late (12–72 h) response phases, each characterized by unique co-regulated gene sets
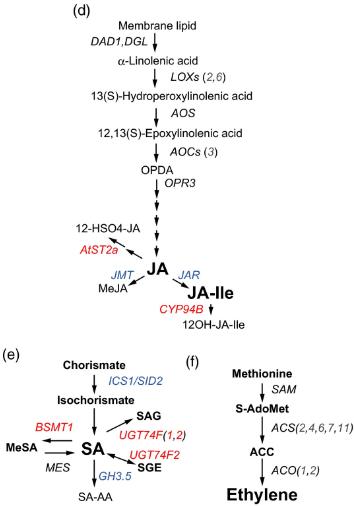
- Coordinated JA–ABA signaling: Dehydration-induced JA accumulation preceded SA changes, and JA-responsive genes showed significant overlap with ABA-responsive transcripts, revealing intimate crosstalk between stress signaling pathways
- AP2/ERF transcription factors identified as early dehydration response regulators, linking hormone signaling to transcriptional reprogramming
- Touch stress-responsive genes distinguished from dehydration-specific responses through the time-resolved experimental design
Process Insight
This study demonstrates that single-hormone (ABA-only) measurements would have revealed the biphasic pattern but missed the JA–ABA crosstalk and the differential regulation of auxin, CK, and GA pathways associated with growth suppression under stress. Comprehensive multi-hormone profiling using LC-ESI-MS/MS with isotope-labeled internal standards was essential to build a systems-level understanding of dehydration responses—an insight directly applicable to engineering drought-tolerant crops.
Where Our Phytohormone Panel Fits
While this study used a research-grade hormone profiling approach, Creative Proteomics supports similar plant stress research by providing:
- Comprehensive phytohormone panels covering auxins, CKs, GAs, ABA, JAs, SA, ACC, BRs, and SLs—all from a single sample submission
- LC–MS/MS and GC–MS/MS (MRM) with isotope-dilution for absolute quantification and cross-batch comparability across multi-timepoint studies
- Pathway-linked outputs (KEGG IDs) and optional multi-omics integration (untargeted metabolomics, transcriptomics, proteomics) for mechanistic insights into hormone-regulated networks
Reference
- Urano, K., Maruyama, K., Jikumaru, Y., Kamiya, Y., Yamaguchi-Shinozaki, K., & Shinozaki, K. (2017). Analysis of plant hormone profiles in response to moderate dehydration stress. The Plant Journal, 90(1), 17–36. https://doi.org/10.1111/tpj.13472
Overexpression of maize ZmLOX6 in Arabidopsis thaliana enhances damage-induced pentyl leaf volatile emissions that affect plant growth and interaction with aphids
Tolley, J. P., Gorman, Z., Lei, J., Yeo, I. C., Nagashima, Y., Joshi, V., ... & Koiwa, H.
Journal: Journal of Experimental Botany
Year: 2023
DOI: https://doi.org/10.1093/jxb/erac522
WI12 Rhg1 interacts with DELLAs and mediates soybean cyst nematode resistance through hormone pathways
Dong, J., & Hudson, M. E.
Journal: Plant Biotechnology Journal
Year: 2022
DOI: https://doi.org/10.1111/pbi.13709
Characterization of CYCLOPHILLIN38 shows that a photosynthesis-derived systemic signal controls lateral root emergence
Duan, L., Pérez-Ruiz, J. M., Cejudo, F. J., & Dinneny, J. R.
Journal: Plant Physiology
Year: 2021
DOI: https://doi.org/10.1093/plphys/kiaa032
Detailed analysis of agro-industrial byproducts/wastes to enable efficient sorting for various agro-industrial applications
Priyanka, G., et al.
Journal: Bioresources and Bioprocessing
Year: 2024
DOI: https://doi.org/10.1186/s40643-024-00763-7
Plant Growth Promotion, Phytohormone Production and Genomics of the Rhizosphere-Associated Microalga, Micractinium rhizosphaerae sp.
Quintas-Nunes, F., Brandão, P. R., Barreto Crespo, M. T., Glick, B. R., & Nascimento, F. X.
Journal: Plants
Year: 2023
DOI: https://doi.org/10.3390/plants12030651
Comparative metabolite profiling of salt sensitive Oryza sativa and the halophytic wild rice Oryza coarctata under salt stress
Tamanna, N., Mojumder, A., Azim, T., Iqbal, M. I., Alam, M. N. U., Rahman, A., & Seraj, Z. I.
Journal: Plant‐Environment Interactions
Year: 2024
DOI: https://doi.org/10.1002/pei3.10155
Physiological, transcriptomic and metabolomic insights of three extremophyte woody species living in the multi-stress environment of the Atacama Desert
Gajardo, H. A., Morales, M., Larama, G., Luengo-Escobar, A., López, D., Machado, M., ... & Bravo, L. A.
Journal: Planta
Year: 2024
DOI: https://doi.org/10.1007/s00425-024-04484-1
Contextualized Metabolic Modelling Revealed Factors Affecting Isoflavone Accumulation in Soybean Seeds
Contador, C. A., et al.
Journal: Plant, Cell & Environment
Year: 2024
DOI: https://doi.org/10.1111/pce.15140
Water-soluble saponins accumulate in drought-stressed switchgrass and may inhibit yeast growth during bioethanol production
Chipkar, S., Smith, K., Whelan, E. M., et al.
Journal: Biotechnology for Biofuels and Bioproducts
Year: 2022
DOI: https://doi.org/10.1186/s13068-022-02213-y
A Water Solution from the Seeds, Seedlings and Young Plants of the Corn Cockle (Agrostemma githago) Showed Plant-Growth Regulator Efficiency
Ambrožič-Dolinšek, J., et al.
Journal: Plants
Year: 2025
DOI: https://doi.org/10.3390/plants14152349